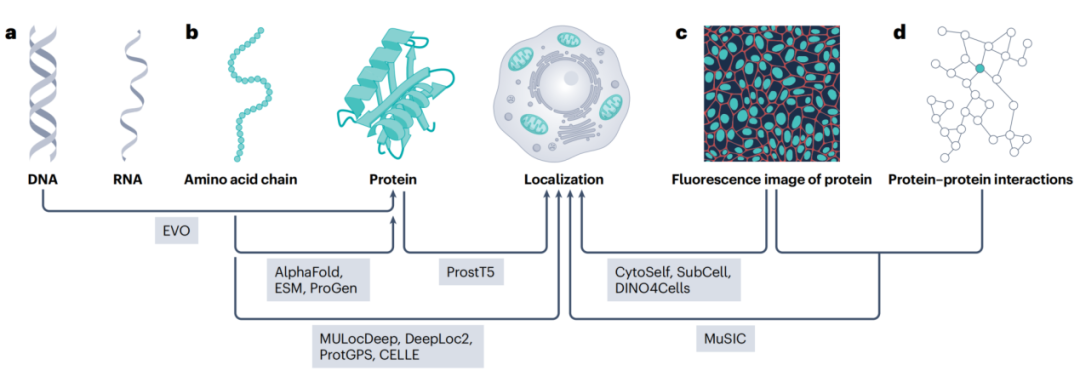
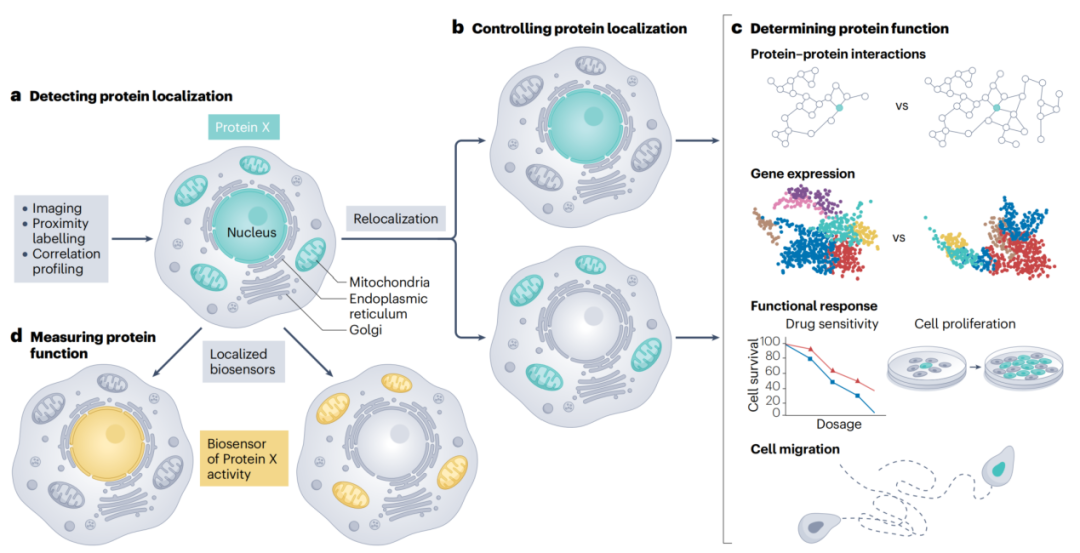
超过一半的人类蛋白会在细胞内"多点办公",这项新技术正在改写我们对蛋白质功能的理解。我们常常把蛋白质想象成在细胞内固定位置工作的"工人":这个蛋白在细胞核里负责转录,那个蛋白在线粒体里负责能量代谢。但真实情况远比这复杂。超过50%的人类蛋白,其实会在细胞内的多个不同位置"多点办公"。核内蛋白偶尔出现在细胞质、细胞膜上的受体也会跑到内体中去。这种"多点分布"不仅普遍存在,更对细胞功能至关重要。
2026年发表在Nature Reviews Molecular Cell Biology(影响因子大于90)上的一篇重磅综述,系统梳理了当前测量和操控蛋白质亚细胞定位的技术全景。今天我们就来解读其中的关键内容。 简单来说,一个蛋白在不同位置,可能执行完全不同的功能。 举个例子:同一个蛋白,当它在细胞膜上时,可能作为信号受体;而当它进入细胞核后,可能直接参与基因调控。如果我们只研究它在某一个位置的功能,就会丢失重要的生物学信息。 更关键的是,很多疾病与蛋白质的错误定位直接相关。比如:
因此,搞清楚蛋白的"行踪"及其调控方式,不仅能帮我们理解细胞运作的基本原理,更可能为疾病治疗提供新靶点。
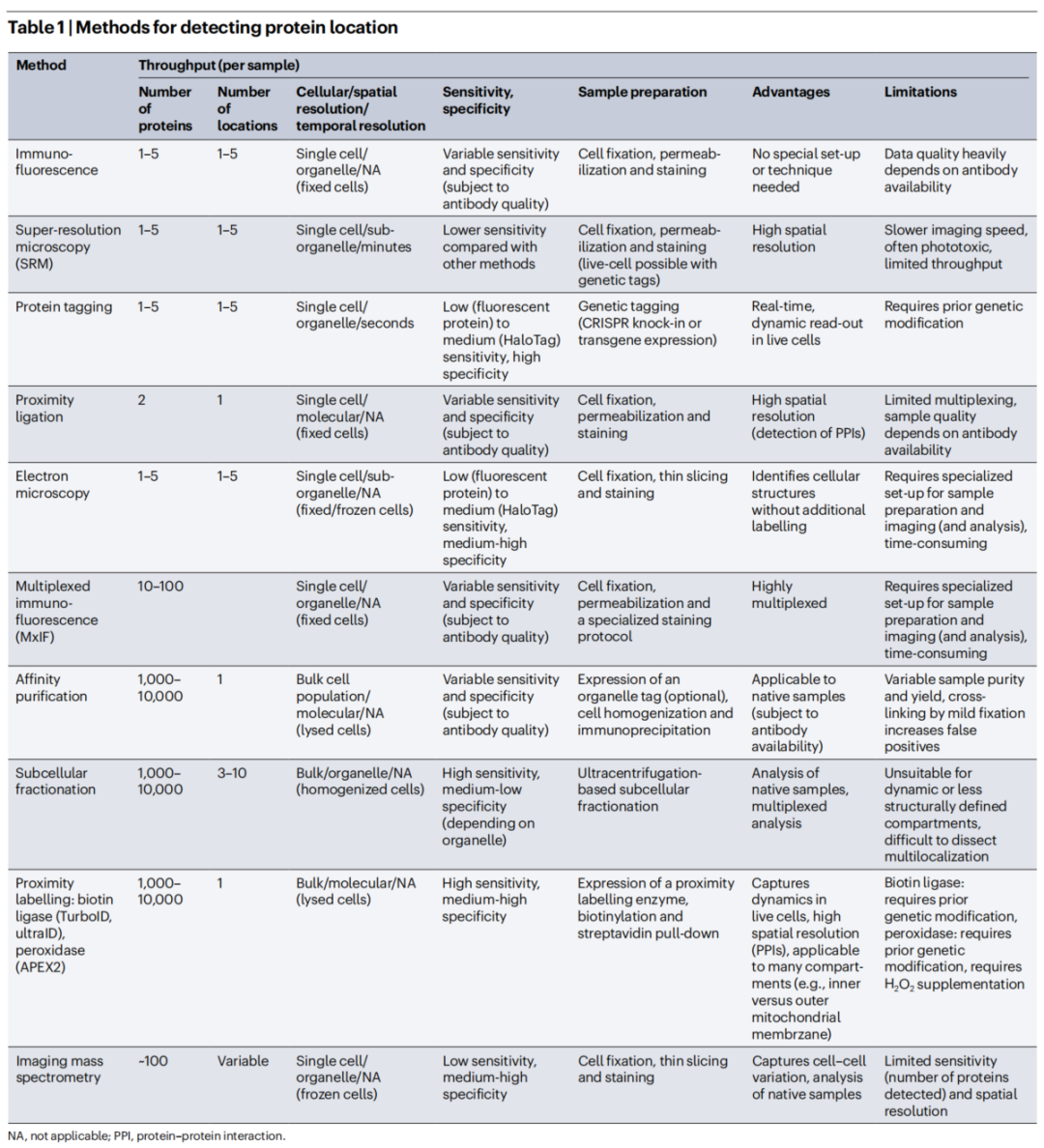
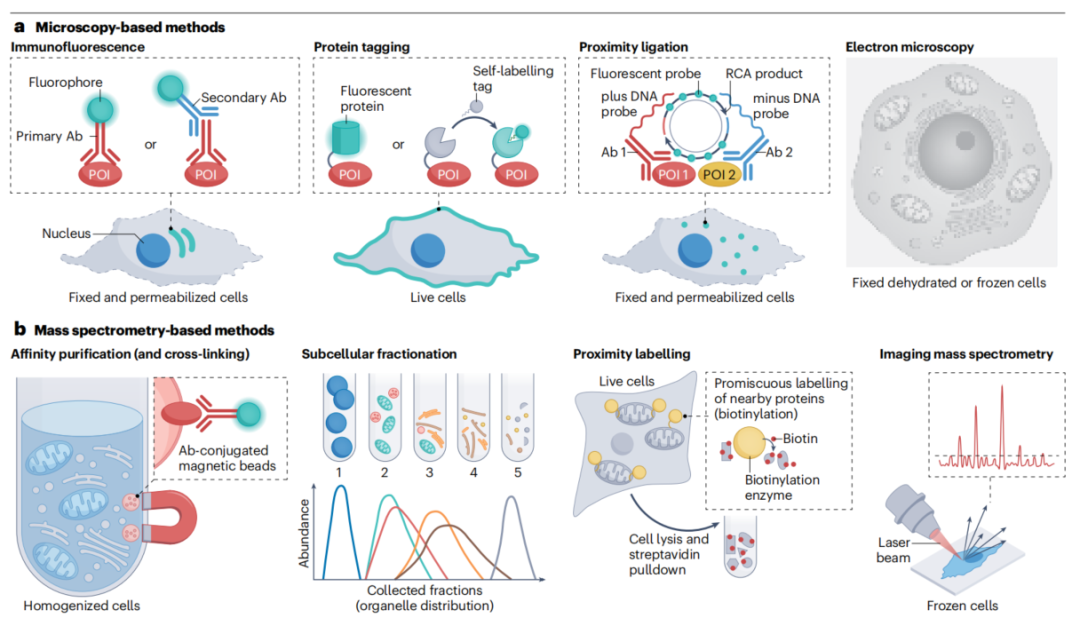
图:模拟细胞蛋白质组的组织结构 综述将测量工具分为两大类:基于显微镜的方法和基于质谱的方法。没有一种方法是完美的,不同方法更多是互补关系。 显微镜类方法 (1)免疫荧光(Immunofluorescence) 这是最基础、最常用的方法。用荧光标记的抗体去标记固定细胞中的目标蛋白,不仅能看到蛋白定位,还能做高通量分析。 但主要瓶颈在于:优质抗体的可及性。商用抗体的失败率可能远超50%,这直接影响了结果的可靠性。 (2)超分辨率显微镜(SRM) 传统光学显微镜的分辨率受限于光波长,只能精确区分相距250纳米以上的两个点。而超分辨率显微镜能突破这个物理极限,实现20-100纳米甚至更高的分辨率。 主要种类包括:
超分辨率的代价是通常成像速度较慢、光毒性较强、通量有限。 (3)蛋白标签技术(Protein Tagging) 为了让蛋白在活细胞中可视化,可以用基因编辑技术在目标蛋白上融合荧光标签。HaloTag、SNAP-tag等自标记蛋白标签,还可以与多种有机染料、生物素等分子结合,灵活性和可调性更强。 (4)邻近连接技术(Proximity Ligation Assay, PLA) 这是我们之前深度讨论过的技术。它的独特优势在于,能将蛋白质空间邻近关系转化为可定量的荧光信号,实现分子水平(<40纳米)的共定位检测。 综述特别提到了PLA的最新升级版——分子像素化(Molecular Pixelation, MPX)。这个新技术不再靠显微镜成像,而是通过DNA测序读取蛋白位置信息,已能同时解析细胞表面超过70种蛋白的空间排布。 (5)电子显微镜(Electron Microscopy) 电子显微镜的分辨率远超光学显微镜,是亚细胞器结构分析的理想工具。通过免疫金标记,可实现约15纳米的蛋白定位精度。但电子显微镜的操作和样本制备都更为复杂,目前在系统性地大规模解析蛋白定位上仍面临诸多挑战。 质谱类方法 (6)亲和纯化质谱(AP-MS) 用"诱饵蛋白"去拉下和它相互作用的"猎物蛋白",然后上质谱鉴定。这个方法能构建大规模的蛋白互作网络,但缺点是容易漏掉瞬时或弱的相互作用。 (7)亚细胞分级偶联质谱(Subcellular Fractionation-MS) 通过差速离心或密度梯度离心,把不同细胞器"分层"分出来,再分别上质谱分析。这个方法曾帮助发现溶酶体,但难以适用于结构模糊或不连续的细胞器(如无膜细胞器)。 (8)邻近标记蛋白质组学(Proximity Labeling Proteomics) 这是近年来的一种热门技术。在目标位置表达一个"生物素轰炸机"(如TurboID或APEX2),它会不加区别地给周围所有蛋白挂上生物素标签。随后用链霉亲和素富集这些"被标记的邻居",再用质谱精确鉴定。 这个技术的威力在于:能在活细胞中快照式地捕获空间蛋白组信息。 更进一步,综述还介绍了动态追踪蛋白搬家过程的方法(TransitID/OrthoID),以及通过光照精准控制标记时间的光控酶(LOV-Turbo)。 表 用于测量蛋白质多定位的工具
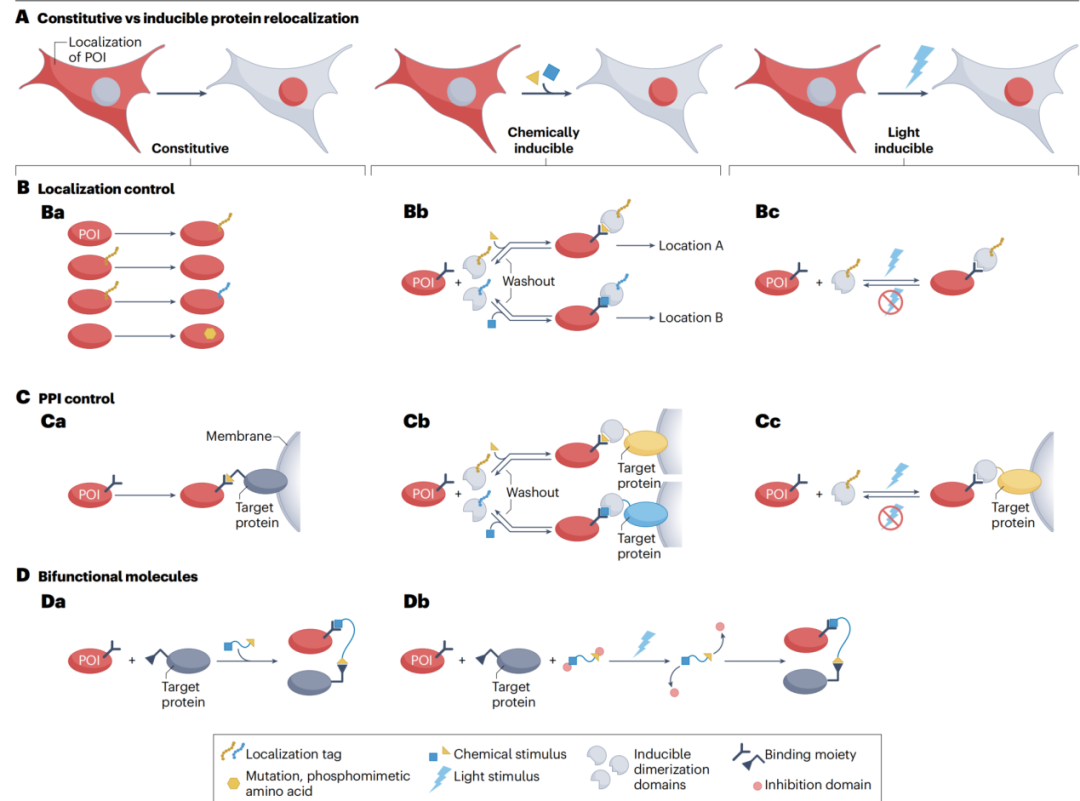
图:蛋白质定位检测方法。 知道了蛋白在哪里还不够,要验证它的定位与功能是否有因果关系,就需要主动操控。 (1)定位标签(Localization Tags) 在目标蛋白上加一段特定的序列,强制让它"搬家"。比如给一个蛋白加上核定位序列,它就会被强行送入细胞核。这种方法适合构建"功能获得"模型,但如果想让蛋白"搬家"并观察其影响,则更适合使用可诱导的瞬时定位技术。 (2)化学诱导的蛋白二聚化(CID) 最经典的是FKBP-FRB-雷帕霉素系统。雷帕霉素像"双面胶"一样,把两个分别融合了FKBP和FRB结构域的蛋白粘在一起。如果把其中一个蛋白锚定在特定细胞器上,另一个蛋白就会被招过去。加药即搬家,洗掉即复原,可逆性很好。 (3)光控蛋白移位(Light-Induced Dimerization, LID) 用光来代替化学药物。与化学诱导相比,光控的时空精度更高、可逆性更好。常用系统如CRY2-CIBN、Magnets等,能被蓝光开启、在暗中关闭。 (4)双功能分子(Bifunctional Molecules) 这类分子可以直接结合两个靶标:一端是"穿梭蛋白"(强烈定位在某个细胞器),另一端是目标蛋白。不需要事先改造目标蛋白,直接"牵手搬家"。如TRAMs(靶向重定位激活分子),甚至已展现临床转化潜力。
图:调控蛋白质定位。
图:确定位置依赖性蛋白质功能的策略
综述还讨论了几个极具潜力的新方向: (1)人工智能预测蛋白定位 AlphaFold等工具能精确预测蛋白的氨基酸序列和三维结构。进一步地,基于这些序列和结构特征,DeepLoc2等模型已能预测蛋白可能去往哪些细胞器,对深层理解蛋白"多点办公"的生物学意义至关重要。 (2)大规模基因编辑筛选 CRISPR技术结合高通量成像,可以系统性地找到"谁在调控某个蛋白的定位"。这在发现疾病中驱动蛋白错位的分子机制方面表现突出,为精准医学提供了可能的干预靶点。 (3)PROTACs的临床转化 PROTACs(蛋白水解靶向嵌合体)是一种可以直接降解特定蛋白的双功能分子。综述指出它正在进入临床试验,有望将蛋白错位相关疾病的机制研究成果,转化为真正的靶向治疗药物。 探秘微观,引领生命科技前沿!精准偶联,开启分子操作新时代;超敏检测,赋能临床检验新应用!杭州乐为生医科技有限公司,精心打造多款科研利器:
选择乐为生医产品,解锁科研新可能。
技术支持:丨13738087339 销售咨询:丨19957893538 欢迎扫描二维码添加微信或拨打电话咨询。 |

 /3
/3 

 浙公网安备33010802005999号
浙公网安备33010802005999号